
سرطان یکی از عوامل اصلی مرگ در سراسر جهان است که در دهههای گذشته پیشرفت قابل توجهی در زمینه تشخیص و درمان آن صورت گرفته است. انواع درمانهای رایج سرطان عبارتاند از پرتودرمانی، شیمیدرمانی و ترکیب آنها که میتوانند علاوه بر حذف سلولهای سرطانی به ایجاد مقاومت، سمیت و صدمه به سلولهای سالم منجر شوند. به طور کلی امروزه پیداکردن یک راه درمانی مناسب، ارزان و با عوارض کمتر برای سرطانهای مختلف یکی از دغدغههای اصلی تحقیقات در زمینه سرطان است.
یکی از این راهکارها، استفاده از پپتیدهای ضدسرطان است که با توجه به انتخابیبودن نسبت به سلولهای سرطانی، عوارض کمتر و اثربخشی آنها مورد توجه بسیاری از دانشمندان قرار گرفته است. یکی از انواع پپتیدهای مورداستفاده، پپتید ضدمیکروبی است که بخشی از پاسخ ایمنی ذاتی در برابر میکروبها، در بسیاری از گونههاست. این پپتیدها، دارای وزن مولکولی کم (40-10 اسیدآمینه) و ساختار آمفیپاتیک و کاتیونی هستند که به آنها اجازه میدهد تا غشاهای منفی سلولهای سرطانی (همانند باکتریها) را هدف قرار دهند. در دهههای اخیر استفاده از این پپتیدها به عنوان عوامل ضدسرطان به عنوان یک روش درمانی نوین در نظر گرفته شده است. پپتیدهای ضدمیکروبی میتوانند از طریق مکانیسمهای مختلفی مانند آپوپتوز، نکروز، لیز غشای سلولی، مهارکردن رگزایی و فعالکردن سیستم ایمنی بر علیه سلول سرطانی فعالیت کنند.
در این مقاله مروری، گروه جدیدی از عوامل ضدسرطان بررسی میشوند که با ساختار پپتیدی، برای بافت توموری دارای اختصاصیت هستند و احتمال مقاومت بدن در برابر سرطان را کاهش میدهند و سمیت کمتری برای سلولهای نرمال بدن دارند؛ لذا این پپتیدها به عنوان یک عامل ضدسرطان، ویژگی هدف قراردادن و آسیبرساندن به غشای سلولهای سرطانی را بدون آسیبرساندن به سلولهای غیرسرطانی و نرمال دارند. از نگاه ساختاری، AMPها یا پپتیدهای ضدمیکروبی دارای دو ساختار شناختهشده آلفا هلیکال و β-sheet هستند. در پایگاه دادههای AMP، بیش از صد نوع AMP با عنوان فعالیت بالقوه ضدتوموری مشخص شدهاند که برای مشاهده لیست کامل آنها میتوان به پایگاه http://aps.unmc.edu/AP/main.php مراجعه کرد. در ادامه نتایج کاربرد برخی از انواع پپتیدهای ضدمیکروبی با فعالیت ضدسرطانی را بررسی خواهیم کرد (جدول شماره 1).
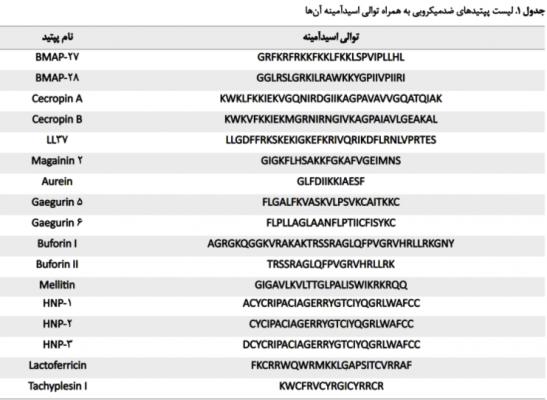
پپتیدهای ضدسرطانی آلفا هلیکال
LL-37
پپتید LL-37 به عنوان یک عامل ضدمیکروبی مشتقشده از کاتلیسیدین انسانی است و نقشهایی نظیر تنظیم پاسخ التهابی، عاملی برای کموتاکسی سلولهای سیستم ایمنی به مکانهای زخم یا عفونت و تحریک رگزایی را به این پپتید نسبت دادهاند. مطالعات متعددی نشان میدهد LL-37 و آنالوگهای آن، علاوه بر اینکه باعث افزایش سمیت در سلولهای سرطانی میشوند؛ در پیشرفت سرطان و متاستاز مؤثرند. بنابراین در مطالعه هیلبورن و همکاران، دریافتند بیان ترانس ژنیک LL-37 به طور قابل توجهی باعث افزایش پرولیفراسیون در سلولهای کراتینوسیت انسان (HaCaT و HEK293) میشود. همچنین یک پپتید کوتاهشده از ناحیه آمین LL-37، به نام LL-25، سیگنالینگ LL-37 را مهار میکند و باعث تغییراتی در مورفولوژی کلونی سلولهای سرطانی میشود. علاوه بر این، وون حوسن و همکاران دریافتند LL-37 در سلولهای سرطانی ریه انسان بیان شده و به عنوان عامل رشد عمل میکند.
قسمتی ازC ترمینال LL-37 (HCAP18: 109 – 135) باعث آپوپتوزیس از طریق دپلاریزاسیون غشای میتوکندری و قطعهقطعه شدن DNA میشود و خاصیت ضدتکثیری روی بسیاری از سرطانها مانند کارسینوم سلول سنگفرشی انسانی و سل لاین SAS-H1 از خود نشان میدهد. پژوهشهای دیگر بیانگر این است که LL-37 از طریق فعالسازی سیگنالینگ به وسیله مکانیسم وابسته پروتئازومها، باعث جلوگیری از تکثیر سلولی در سرطان معده میشود. همچنین این پپتید میتواند باعث ایجاد فرایند آپوپتوزیس از طریق مسیر مرتبط با میتوکندری در سلولهای لوسمی انسانی jurkat شود. FK-16، که یک قطعه کوتاهتر از LL-37 است، میتواند باعث آپوپتوزیس مستقل ازکاسپازها و اتوفاژی از طریق مسیر p53-Bcl 2/Bax در سلولهای سرطانی روده بزرگ شود.
بر خلاف AMPهای دیگر، LL-37 برای سلولهای یوکاریوتی در غلظتهای 30-25 میلیمولار که 3 تا 5 برابر مقدار MIC آن است، سمی است.LL-37 میتواند به غشاهای لیپیدی متصل شود و سپس باعث تغییر شکل در سر گروه فسفولیپیدی آن شده و به ایجاد فشار در دو لایه لیپیدی منجر و درنهایت باعث ایجاد یک اختلال در قسمت هیدروفوبیکی غشا شود.
همچنین این پپتید میتواند به عنوان یک شمشیر دولبه عمل کند؛ به گونهای که در بعضی از انواع سرطان با آن مقابله میکند و در عین حال میتواند باعث وخیمترشدن آن و افزایش تکثیر سلولهای سرطانی شود. تمامی این مکانیسمها بستگی به این دارد که LL-37 از چه طریقی با غشای سلول ارتباط برقرار میکند.
این پپتید زمانی که با گیرنده EGFR برخورد کند میتواند باعث افزایش تکثیر سلولی وکاهش پاسخ ایمنی ذاتی شود. همچنین از طریق برخورد با گیرندههای G-protein coupled receptor و FRP2 یا FRPL1 میتواند محرک فرایند آنژیوژنزیس و کموتاکسی شود. بر اساس این یافتهها، شگفتآور نیست که LL-37 با پیشرفت سرطان و متاستاز مرتبط است. درواقع، بیان hCAP18 / LL-37 در سلولهای سرطانی سینه افزایش مییابد و تولید آن در ناحیه اپیتلیوم سرطانهای بدخیم سینه، بیشتر از اپیتلیالهای معمولی یا تومورهای خوشخیم است. همچنین یک پپتید کوتاهشده از ناحیه آمین LL-37، به نام LL-25، سیگنالینگ LL-37 را مهار میکند و باعث تغییراتی در مورفولوژی کلونی سلولهای سرطانی میشود.
بنابراین میتوان گفت این پپتید یک هدف درمانی قابل پیشگیری برای جلوگیری از پیشرفت بیماری متاستاتیک در بعضی از انواع سرطان است، اگرچه مکانیسمهای دقیق مولکولی آن هنوز به طور کامل مشخص نشده است. LL-37 در سرطانهای پستان، ریه و پروستات باعث گسترش، تکثیر، مهاجرت و تومورزایی از طریق دریافت سیگنال گیرندهها میشود، در حالی که در سرطان معده، روده بزرگ و سلولهای T، میتواند باعث توقف گسترش و منجر به فرایند مرگ سلولی یعنی آپوپتوز و اتوفاژی شود.
ملیتین
ملیتین، یک پلیپپتید محلول در آب، خطی، کاتیونی، همولیتیک و آمفیپاتیک و شامل 26 اسیدآمینه است و به عنوان مولکول اصلی فارماکولوژیک زهر نیش زنبور،40 تا 50 درصد از وزن خشک آن را تشکیل میدهد. ناحیه N ترمینال آن عمدتاً دارای آمینواسیدهای هیدروفوبی و انتهای C ترمینال آن حاوی آمینواسیدهای مثبت و هیدروفیلی است. ملیتین میتواند به سطح غشای منفی سلولهای سرطانی متصل شده و سپس کل دو لایه فسفولیپید غشا را با تشکیل حفرهها مختل کند. این امر همراه با نشت یونهای اتمی و مولکولها و افزایش نفوذپذیری غشاست که درنهایت به لیزشدن سلول منجر میشود. از مکانیسمهای سایتوتوکسیک ملیتین برای ازبینبردن سلولهای سرطانی میتوان به القای آپوپتوزیس، مکانیسمهای جلوگیری از متاستاز و تهاجم سلولهای سرطانی، ایجاد تغییر در سیکلهای سلولی و تأثیر بر رگزایی تومور اشاره کرد.
به علت فقدان نسبی انتخابیبودن ملیتین برای سلولهای سرطانی، تلاشها برای هدفدارکردن ملیتین بدینگونه است که از ماتریکس متالوپروتئیناز-۲ که در سلولهای سرطانی انسان و اندوتلیوم مرتبط با تومور، بیش از حد بیان میشود، استفاده شده است. سلولهای کارسینوم DU 145و سلولهای کارسینوم تخمدان SKOV3 که سطح بالایی از ماتریکس متالوپروتئیناز-۲ را بروز میدهند، توسط ملیتین آویدین کونژوگهشده، کشته میشوند؛ این در حالی است که فیبروبلاستها که دارای ماتریس متالوپروتئیناز-۲ کمتری هستند، کمتر تحت تأثیر اثرات سایتوتوکسیک این پپتید کونژوگه قرار میگیرند. علاوه بر این، تزریق داخل توموری ملیتین آویدین کونژوگه موجب کاهش قابل توجهی در رشد سلولهای ملانوم B16 در موش سینژنیک میشود. همچنین آنتیبادیهای اختصاصی تومور میتوانند برای هدف قراردادن ملیتین برای سلولهای تومور استفاده شوند. ملیتین همچنین میتواند اثرات بیشتری روی سلولهای سرطانی که دچار افزایش بیان Ras شدهاند داشته باشد که این کار از طریق مکانیسمهایی شامل فعالسازی بیش از حد فسفولیپاز A2، نفوذ کلسیم و تخریب سلولهای تغییر شکل یافتهشده انجام میشود.
همچنین نتایج نشان میدهد اثر مهاری ملیتین روی مهاجرت سلولهای سرطان سینه ممکن است به مهار مسیر mTOR مرتبط باشد. پژوهشی که در سال 2016 روی مکانیسم آپوپتوز این پپتید برای درمان سرطان معده انجام شد، مشخص کرد سل لاین SGC-7901 که در معرض ملیتین قرار دارد نسبت به گروه کنترل، مقدار سیتوکروم C و پروتئینهای AIF افزایش یافته و سطح Smac/diablo کاهش مییابد. در سال 2014 در مطالعه دیگری، اثرات مهاری ملیتین روی متاستاز و تهاجم القاشده توسط EGF سلولهای سرطان سینه تأیید شد. علاوه بر این، ملیتین باعث مهار بیان MMP9 القاشده توسط EGF از طریق مسدودکردن مسیرهای NF-κB ،PI3K ،Akt و mTOR در این سلولها میشود و همچنین شدت فسفوریلاسیون FAK را کاهش میدهد.
در سال 2014 در تحقیق دیگری که درباره مکانیسم تأثیر ملیتین روی سیکلهای سلولی در سرطان هپاتوسلولار انجام شد، متوجه شدند MEL مانع از تکثیر سلولی میشود؛ به طوری که CyclinD1 و CDK4 را به طور قابل توجهی کاهش میدهد. علاوه بر این، ملیتین قادر به افزایش بیان PTEN که یک نوع ژن سرکوبگر سرطان است، شده و در عین حال باعث تضعیف بیان هیستون داستیلاز میشود. درمان با ملیتین موجب کاهش فسفوریلاسیون Akt میشود.
BMAP
پپتیدهای BMAP در دو نوع BMAP-27 وBMAP-28 با توالیهای ۲7 و ۲8 آمینواسیدی از کاتلیسیدین گاوی استخراج شدهاند. انتهای آمین این پپتیدها خاصیت کاتیونی دارد و به نظر میرسد یک ساختار آلفا هلیکس آمفیپاتیک را تشکیل دادهاند و دم هیدروفوبی این پپتیدها برای خاصیت سایتوتوکسیستی آن ضروری است. فعالیت سایتوتوکسیتی BMAP-27 و BMAP-28 بر سلولهای نئوپلاستیک نشان داد غلظت 5/1 تا 6 میکرومولار از پپتید BMAP، منجر به افزایش نفوذپذیری غشا، ورود یون کلسیم و قطعهقطعهشدن DNA و درنهایت موجب آپوپتوز سلول میشود.
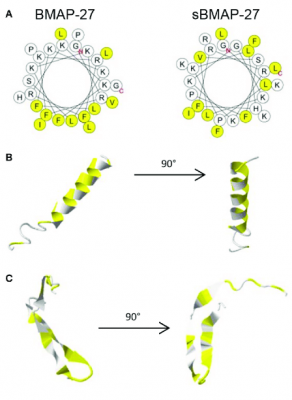
همچنین درمان با BMAP-28 در سلولهای سرطانی انسانی U937 و K562، باعث بازشدن منافذی در غشای میتوکندری و کاهش سریع پتانسیل غشای این سلولها میشود که درنهایت با آزادشدن سیتوکروم c به شروع مرگ سلولی یا آپوپتوز منجر میشود. دادهها حاکی از آن است که BMAP-28 به طور قابل ملاحظهای از تکثیر سلولهای سرطان تیروئید در شرایط برونتنی جلوگیری میکند و باعث اثرات آپوپتوزی در این سلول سرطانیشده و با افزایش بیان کاسپازهای فعال 3 و 9، سطوح رونویسی و ترجمه آن ها را افزایش میدهد و همچنین، بیان ماتریکس متالوپروتئینازهای-3 و 9 تحت درمان با BMAP-28 کاهش مییابد که این امر نشان دهنده کاهش فعالیت متاستاتیک سلولهای سرطانی تحت درمان با این پپتید است.
استفاده از غلظتهای بالاتر این پپتید باعث ایجاد پاسخهای درمانی بهتری نشده و در بعضی موارد شاهد کاهش قابل ملاحظهای از نوتروفیلها و اریتروسیتهای طبیعی بدن انسان هستیم، زیرا با وجود خاصیت سایتوتوکسیک پتیدهای BMAP-27 و BMAP-28 در غلظتهای کم در برابر سلولهای سرطانی؛ در غلظتهای بالا برای لنفوسیتهای فعالشده انسانی نیز دارای خاصیت سمیت هستند.
سکروپین
سکروپین A و B که از همولنف یک مگس ابریشم غولپیکر جدا شده است، ساختاری شامل دو آلفا هلیکس، دارای انتهای آمینآمفی پاتیک و انتهای کربوکسیلی هیدروفوبیک، با فعالیت سیتوتوکسیک ضدسرطان مرتبط با انتهای آمین هستند. سکروپین A و B میتوانند انواع مختلفی از سلولهای سرطانی انسان را در غلظتهای مختلف که برای سلولهای طبیعی یوکاریوتی مضر نیستند، لیز کنند. تأثیر سکروپین B در سلول کارسینوم معده انسانی، منجر به کاهش ایجاد منافذ کانالهای غشای گذر میشود. سکروپین B ممکن است برای درمان سرطانهای انسانی پتانسیل زیادی داشته باشد، زیرا این پپتید فعالیتهای ضدتوموری در موشهای حاوی سلولهای آدنوکارسینوما کولون و همچنین فعالیتهای سیتوتوکسیک در برابر سلولهای سرطانی تخمدان مقاوم به دارو را از خود نشان داده است.
ترکیب سکروپین A و عوامل شیمیدرمانی معمولی مانند 5-فلورویوراسیل و سیتارابین در غلظتهای خاصی، اثرات سیتوتوکسیسیتی سینرژیک را روی سلولهای لوسمی لنفوبلاستی انسان CCRF-SB از خود نشان میدهند. پپتید سکروپین-B به ایجاد کانال در غشای سلولهای سرطانی منجر میشود؛ در حالی که آنالوگ سکروپین-B3 که متشکل از دو رشته آلفا هلیکس هیدروفوب است، موفق به ایجاد حفره در غشا نمیشود. همچنین آنالوگ سکروپین-B1 که دارای دو رشته آمفیاتیک آلفا هلیکس است، فعالیت سیتوتوکسیک قویتری در برابر چندین رده از سلولهای سرطانی لوسمی انسانی در غلظتهای مختلف از خود نشان داده، اما تأثیری بر فیبروبلاستهای طبیعی یا اریتروسیتهای طبیعی انسان نداشتند.
این یافته ها نشان میدهد انتهای آمین آلفا هلیکس با ویژگی آمفیپاتیک سکروپین-B میتواند با استفاده از قسمتهای اسیدآمینهای و بازی خود، با مولکولهای غشایی آنیونی ارتباط برقرار کند و فعالیت سیتوتوکسیک خود را در برابر سلولهای سرطانی با این روش ایجاد کند، در حالی که انتهای کربوکسی آن چنین عملکردی ندارد. با وجود این، C ترمینال آلفا هلیکس این پپتید ممکن است نفوذپذیری غشا توسط این پپتیدها را افزایش دهد. در مطالعات اخیری که روی این پپتید انجام شده است، سمیت سکروپین A و آنالوگ آن (ABP-dHC-Cecropin A-K) روی سه عدد از سللاینهای سرطان لوکمی و دو عدد از سللاینهای غیرسرطانی به اثبات رسید.
مگینین
مگینین به عنوان یک ماده سایتوتوسیک اختصاصی برای سلولهای سرطانی، از پوست قورباغه پنجهای آفریقایی جدا شده است. مگینین شامل ۲1 تا ۲7 آمینواسید با یک ساختار ثانویه آلفاهلیکس است که ساختار کاتیونی و هیدروفوبی دارد. یکی از آنالوگهای این پپتید مگینین-2 است. مگینین-2 و آنالوگهای قویتر آن، شامل مگینین A ،B و G هستند که به علت ایجاد کانالهای یونی α-Helical به آسیب به غشای سلولهای سرطانی منجر میشوند. فرایند اختصاصیت بالا برای غشای سلولهای سرطانی توسط مگینین-G کشف شده است، در حالی که مگینین-B قویترین آنالوگ سنتتیک از نظر سایتوتوکسیسیتی است. این یافتهها نشان میدهد مگینین-2 در برابر سلولهای سرطانی دارای سیتوتوکسیسیتی اختصاصی است و میتواند باعث ایجاد آپوپتوز شود. مگینین-2 ممکن است یک شیوه درمانی جدید و مؤثر در درمان سرطان کولون با اثرات بسیار کم سیتوتوکسیک برای سلولهای طبیعی باشد.
مگنین-2 و آنالوگهای آن میتوانند دو رده از سلولهای سرطانی شامل هماتوپوئیتیک و تومورهای بافتی را در غلظتهای 5 تا 10 برابر کمتر از غلظتهای مؤثر مگینین که برای لنفوسیتهای خون محیطی انسان یا نوتروفیلها سمی هستند، بهسرعت لیز کنند. مگینین-2 همچنین فعالیتهای سیتوتوکسیک اختصاصی در برابر چندین رده از سلولهای سرطانی شامل سلولهای سرطانی مثانه در انسان با میانگین IC50 تقریباً ۲00 میکرومولار را از خود نشان میدهد. مگینین A و مگینین G مانع رشد سلولهای کوچک سرطانی ریه انسانی، از جمله انواع سلولهای تومور مقاوم به دارو، با غلظت IC50 تقریباً 9 میکرومولار میشو. در مطالعهای که اخیراً در سال 2016 درباره درمان سرطان کلورکتال انجام شد، مگینین-2 باعث جلوگیری از زندهماندن رده سللاین سرطانی colo320 DM در غلظت 89/3 نانومولار شد.
آئورین
پپتیدهای آئورین یک خانواده پپتیدی کشفشده در ترشحات پوستی قورباغههای استرالیایی هستند که با درنظرگرفتن طول و سایزشان طبقهبندی میشوند. پپتید آئورین 1/2 یکی از کوتاهترین پپتیدها در این خانواده با 13 آمینواسید است که از نظر ساختاری، دارای خاصیت آمفیپاتیک است که با قسمتهای هیدروفوب و هیدروفیلی، در امتداد محور مارپیچ آلفا هلیکس آنها تشکیل میشود و یکی از قویترین پپتیدهای ضدمیکروبی است. این پپتید کاتیونی بهآسانی در حضور غشای فسفولیپیدی، به حالت آلفا هلیکس تبدیل میشود و به طور عادی میتواند به غشاهایی با بار معمولی نیز وصل شود؛ این در حالی است که دارای افینیتی بسیار بالاتری نسبت به غشاهای آنیونی مانند غشای سلولهای سرطانی است. معمولاً تعامل این پپتید با غشا، روی سطح آن رخ داده و میتواند باعث بههمریختن یکپارچگی غشا به طور وابسته به غلظت شود. این پپتید، فعالیت ضدسرطانی متوسطی را در برابر 52 مورد از 54 سللاینهای سرطانی مورد تأیید NCI، بدون آسیبرساندن به سلولهای پستانداران از خود نشان داده است. در مطالعات اخیری که در سال 2017 که روی سلولهای MCF-7 با استفاده از پپتیدهای Aurein 1 و 2 انجام شد، مشخص شد این پپتیدها در یک غلظت معین دارای خاصیت لیزکننده غشایی هستند و میتوانند با اثر روی غشای سلولی، باعث خروج کلسئین شوند که یک ماده فلورسانس جهت تشخیص نشت در غشای لیپیدی است.
گاگورین
گاگورین که از پوست یک قورباغه کرهای جدا میشود، دارای یک ساختار رندوم کویل در محلول آبی و یک ساختار آمفیپاتیک آلفاهلیکال در محیطهای غشایی است. گاگورین 5 و 6 هریک از 24 ریشه اسیدآمینهای تشکیل شدهاند و هر دو به عنوان آنالوگ پپتیدی سنتتیک، قادر به انتخاب گزینشی انواع مختلف سلولهای سرطانی انسانی، از جمله HCT116 کولون و سلولهای کارسینوم پستان MCF-7 هستند، در حالی که کمترین فعالیت همولیتیک را از خود نشان میدهند. برخی از مطالعات نشان دادهاند گاگورین-5 و گاگورین-6 دارای فعالیت اختصاصی سیتوتوکسیک در برابر سلولهای نئوپلاستیکی هستند.
گاگورین-6 و آنالوگهای پپتیدی و سنتیتک آن دارای طیف گستردهای از فعالیتهای سیتوتوکسیک در برابر سلولهای سرطانی انسانی همراه با کمترین سمیت سلولی برای سلولهای تکهستهای خون محیطی و حداقل فعالیت همولیتیک هستند. علاوه بر این، گاگورین-6 میتواند از طریق مکانیسم آپوپتوز به عنوان عامل سیتوتوکسیسیتی در برابر سلولهای سرطان پستان MCF-7 باشد، زیرا شواهدی از DNA fragmentation در سلولهای سرطانی که با این پپتید مواجه شده بودند، وجود دارد.
بوفورین
بوفورین-I یک پپتید 39 اسیدآمینهای است که برای اولینبار از بافت معده قورباغه آسیایی جدا شده است. بوفورین-II که از بوفورین-I ساخته میشود، یک پپتید 21 آمینواسیدی است که فعالیت ضدمیکروبی قویتری را نسبت به پپتید اصلی آن، یعنی بوفورین-I نشان میدهد. بوفورینها از طریق انتقال به سلول، بدون افزایش نفوذپذیری در غشا و درنتیجه اتصال به اسیدنوکلئیک میتوانند باعث مرگ سلولی شوند. برخلاف چندین پپتید کاتیونی دیگر، بوفورین-II اثرات سیتوتوکسیک در برابر سلولهای طبیعی یوکاریوتی از خود نشان نمیدهد. به عنوان مثال، بوفورین-II حتی در غلظت بیش از 200 برابر مورد نیاز برای مهار رشد باکتری (MIC)، تقریباً در برابر اریتروسیتهای انسانی فعالیت غیرهمولایتیک دارد. همچنین هنگامی که فعالیت سیتوتوکسیک آن روی سلولهای TM12 فیبروبلاستی انسان اندازهگیری شد، بوفورین-II هیچ تأثیری روی حیات سلولی سلولها در غلظت 100 میکرومولار نداشت.
یک مطالعه دیگر نشان داد بوفورین-II و بوفورینIIb- که یک آنالوگ مصنوعی بوفورین-II است، فعالیت سیتولیتیک اختصاصی در برابر 62 سللاین سرطانی با غلظت پپتیدی IC50 در رنجی حدود 24 μg/mL از خود نشان میدهد. بوفورینIIb- به طوراختصاصی سلولهای سرطانی را از طریق تعامل با گانگلیوزیدهای سطح سلول، هدف قرار میدهد. این اختصاصیت به طور عمده، ناشی از عدم توانایی پپتید برای نفوذ در غشاهای سلولهای طبیعی است. سطح غیرقابل نفوذ غشای سلولهای طبیعی پستانداران، عمدتاً از فسفولیپیدها و استرولهاست و همین امر باعث نفوذناپذیری این غشاها توسط این پپتید میشود. بوفورین-IIb سپس غشای سلول سرطانی را بدون آسیبرساندن به آن متوقف میکند و باعث آپوپتوز وابسته به میتوکندری میشود. این پپتید اثرات سیتوتوکسیک زیادی را هنگام تزریق به تومورهای جامد در موشهای دارای کمبود P53 از خود نشان میدهد. این نتایج نشان میدهد بوفورین-IIb ممکن است یک کورسوی امید نوینی در جهت درمان بسیاری از سرطانها باشد.
پپیتدهای ضدسرطانی دارای ساختار sheet-β
دیفنسین
دیفنسینها گروهی از AMPهای کاتیونی غنی از سیستئین و آرژنین و حاوی 18تا 45 اسیدآمینه هستند. برخی از آنها به عنوان آلفا دیفنسین، توسط نوتروفیل انسانی تولید شده که از HNP-1 تا HNP-4 نامگذاری میشوند یا توسط سلولهای پنت در ایلئوم انسان ساختهشده و به نامهای HD5 و HD-6 نامیده میشوند. دسته دیگری از خانواده بتا دیفنسینهای انسانی شناسایی شدهاندکه شش عضو این خانواده (hBD-1 تا hBD-6) عمدتاً توسط سلولهای اپیتلیال ترشح میشوند.
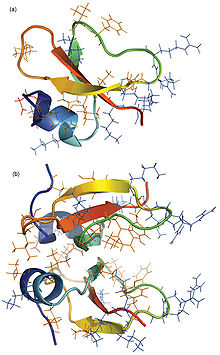
ساختار مونومری و دیمری بتا-دیفنسین HBD-2 انسان
کشتهشدن سلولهای تومور توسط 2، 1HNP- و 3 شامل یک روند اتصال به غشاست که احتمالاً از طریق تعاملات الکترواستاتیک و پس از آن کمشدن سریع پتانسیل غشا و ازبینرفتن یکپارچگی غشا انجام میشود. سایتوتوکسیسیتی وابسته به HNP ممکن است باعث آسیب به DNA شود، زیرا شکستهای DNA تکرشته در سلولهای هدف HNP شناسایی شده است و قطعاتی در اندازههای نوکلوزوم که مشخصه آپوپتوز هستند، مشاهده نشد. عملکرد این پپتید روی حیات، تکثیر و اختلال در سیکلهای سلولهای سرطانی نشان داده است دیفنسین در غلظتهای کمتر به طور قابل توجهی باعث تحریک پرولیفراسیون و زندهماندن سلولهای سرطانی میشود و در غلظتهای بالاتر منجر به مهار قابل توجه، تکثیر و مهاجرت سلولی یا متاستاز میشود.
غلظتهای بالاتر از ۲5 میکروگرم در میلیلیتر از 2، 1HNP- و 3 باعث سرکوب سنتز DNA در سلولهای کارسینوما سلولهای کلیه و همچنین کاهش حیات سلولی سلولهای سرطانی میشود. در غلظتهای 1 تا 100 نانومولار hBD-4 به طور قابل توجهی باعث تحریک پرولیفراسیون و زنده ماندن سلولهای سرطانی میشود و پیشرفت سلولهای سرطانی از طریق نقاط بازرسی G2/M را افزایش میدهد و در کل باعث افزایش فعالیت سلولی میشود. در درمان این سلولها با غلظت 500 نانومولار، rec-hBD-4 منجر به اثرات متقابل شده و مهار قابل توجه تکثیر و حیات سلول، انسداد چرخه سلولی در نقاط بازرسی G1/S، مهار متاستاز و تشکیل کلنی از اثرات این پپتید در غلظت بالاتر است.
اثرات ضدسرطان دیفنسین-3 انسانی (hBD3) و همولوگ آن Defb14، پس از 9 روز تزریق مداوم روی مدل موشی سرطان ریه، به طور قابل توجهی قویتر از فعالیت سایر ایزوفرمهای دیفنسین بود. در سال ۲008 زو و همکارانش نشان دادند تزریق داخل توموری دیفنسین-1 آلفا باعث جلوگیری از رشد سلولهای آدنوکاریسینوم ریه انسان در موشهای ناد شده و باعث آپوپتوز این سلولها میشود.
لاکتوفریسین
لاکتوفریسین یک AMP کاتیونی تولیدشده توسط هیدرولیز لاکتوفرین و یک پروتئین متصلشونده به آهن است که در گرانولهای ترشحی نوتروفیلها و در مایعات مترشحه مانند شیر و بزاق وجود دارد. لاکتوفریسین گاوی یا LfcinB دارای اثرات سیتوتوکسیک در برابر بسیاری از انواع مختلف سلولهای سرطانی موشی و انسانی، از جمله سلولهای لوسمی، سلولهای فیبروسارکوم، کارسینومهای مختلف و سلولهای نوروبلاستوماست. فعالیت سیتوتوکسیک لاکتوفریسین-B در برابر سلولهای سرطانی به میزان زیادی بستگی به ساختار آمفیپاتیک و بار مثبت خالص این پپتیدها دارد.
تحقیقاتی که روی این پپتید انجام شده، نشان میدهد مصرف لاکتوفریسین موجود در شیر گاو در طولانیمدت میتواند تعداد سلولهای تکثیریافته را کاهش دهد و با افزایش این فاز، باعث ازدیاد زمان ترمیم DNA شود و از این طریق از ایجادشدن سلولهای سرطانی جلوگیری کند. همچنین برخی مطالعات نشان میدهد لاکتوفرین گاوی توانایی ضدسرطانی بسیار زیادی از طریق فعالسازی آبشارهای سیگنالینگ، شامل القای پروتئین P53 ، آپوپتوزیس و آنتیآنژیوژنزیس دارد.
در سال 2018 اثرات سیتوتوکسی مستقیم این پپتید روی رده سلولی HNSCC مقاوم و حساس به سیسپلاتین، بررسی شد که با کاهش IL-6 و PD-L1 همراه بود و این یعنی LfcinB ممکن است بر مقاومت در برابر سیسپلاتین غلبه کند. در موشهایی که رشد HNSCC گزنوگرفت مقاوم به سیس پلاتین داشتند،پس از تزریق LfcinB به مدت سه روز در مقایسه با حیوانات گروه کنترل کاهش قابل توجهی در حجم تومور از خود نشان دادند.
در سال 2017 که از مشتقات دایمریک و تترامریک حاوی موتیف RRWQWR از LfcinB روی سللاینهای سرطان سینه یعنی MDA-MB-468 و MDA-MB-231 استفاده کردند، در هر دو مورد سایتوکسیسیتی را روی سلولها ارزیابی کردند و شاهد اثرگذاری این مشتقات در ازبینرفتن این سللاینهای سرطانی بودند.
تاکی پلسین
هوموسیت خرگوش نعل اسبی، منبع این AMP کاتیونی است که شامل 17 اسیدآمینه است. این پپتید دارای صفحات بتا همراه با پیوندهای دیسولفیدی است. این ساختار اجازه میدهد تا تمام شش اسیدآمینه بازی آن، یعنی لایزین و آرژینین در سطح پپتید و در دسترس قرار گرفته و ساختاری آمفیپاتیک را به وجود آورد. تاکیپلسین-1 میتواند در هیالورونان موجود در سلولهای سرطان انسانی که دارای بیان بیش از حد هیالورونان هستند و همچنین به عنوان جزء مکمل C1q سرم در بدن انسان، منجر به فعالشدن مسیرکمپمان کلاسیک و لیزشدن سلول سرطانی شود.
اخیراً چندین مورد مطالعه روی این پپتید صورت گرفته که اهمیت این پپتید در درمان سرطان را نشان میدهد. در سال 2018 درباره اثر ترکیبی پپتیدهای ضدمیکروبی تاکی پلسین-I و سیسپلاتین بر سلولهای تومور و سلولهای طبیعی انسان مطالعه شد. آزمون MTT و فلوسایتومتری نشان داد تاکیپلسین-I به طور انتخابی و اختصاصی سلولهای سرطانی را همراه به سیسپلاتین به طور وابسته به غلظت میتواند تحت تأثیر قرار دهد. آزمایشهایی نشان داد استفاده ترکیبی از تاکیپلسین-I و سیسپلاتین موجب کاهش دُز موثر سایتوکسیک و همچنین کاهش دُز سمیت غیراختصاصی آن میشوند.
در سال 2017 اثرات استیلاسیون N ترمینال و اضافهکردن گروه آمید انتهای C ترمینال، بر ویژگیهای سیتوتوکسیک تاکی پلسین-I بررسی شد و تاکیپلسین-I اصلاحشده در برابر سلولهای تومور و سلولهای طبیعی، سایتوتوکسیسیتی بیشتری از خود نشان داد. فعالیتهای همولیتیک تاکیپلسین-I اصلاحشده نیز بیشتر از مولکول اولیه آن بود. در مقایسه با تاکی پلسین-I اصلاح نشده، این پپتید با اصلاح C و N ترمینال، مقاومت بیشتری در برابر تجزیه پروتئولیتیک در سرم تازه انسان از خود نشان میدهد.